Research Interests
Our research focuses on understanding processes that affect the dynamics of populations. In some cases we focus on processes that may be characterized as 'ecological', e.g., variation in larval settlement, density-dependent mortality, species interactions. In other cases, we focus on processes that may be characterized as 'evolutionary', e.g., understanding heritable genetic variation and natural selection via differential mortality or reproduction. However, much of our research seeks to understand the interactions between ecological and evolutionary processes. Our approach combines empirical research, quantitative analyses, and development of theory in roughly equal proportions. Details about some of our research projects are summarized below.
Current Research Projects:

Demography & population dynamics of kelp forest fishes
Much of the current research in our lab is geared toward understanding the dynamics of nearshore fish populations. Central to this endeavor is understanding variation in the demographic rates that are fundamentally responsible for changes in population size. We measure survival, growth, and reproduction of fishes in the field by using a variety of methods, including visual transect surveys, size comparisons, and mark-recapture studies.
We are interested in how demographic rates vary across multiple spatial scales and how demographic rates are affected by various environmental factors (e.g., predator abundance, habitat complexity, oceanography). We also study variation among individuals and whether particular phenotypes (e.g., growth rates, behavior) are associated with survival and/or reproduction. Such relationships are particularly important in the context of natural selection.
Much of the current research in our lab is geared toward understanding the dynamics of nearshore fish populations. Central to this endeavor is understanding variation in the demographic rates that are fundamentally responsible for changes in population size. We measure survival, growth, and reproduction of fishes in the field by using a variety of methods, including visual transect surveys, size comparisons, and mark-recapture studies.
We are interested in how demographic rates vary across multiple spatial scales and how demographic rates are affected by various environmental factors (e.g., predator abundance, habitat complexity, oceanography). We also study variation among individuals and whether particular phenotypes (e.g., growth rates, behavior) are associated with survival and/or reproduction. Such relationships are particularly important in the context of natural selection.

Black surfperch (Embiotoca jacksoni). Photo credit: Steve Lonhart
|
Special little snowflakes: Why individual variation matters for the dynamics of populations
Much of ecological theory treats individuals as though they are identical. Of course, Biologists know that individuals vary. After all, anyone with a loving mother knows that they are a beautiful, special little snowflake and that there is no one else quite like them. However, Biologists often need to make simplifying assumptions, and one of those assumptions is that variation among individuals does not have significant effects on the dynamics of populations. In other words, we know that individuals vary, but we think we can ignore this when it comes to studying higher-order phenomena (e.g., changes in population size). |
Our recent study (Johnson et al. 2016, Ecology) examined dynamics of black surfperch populations at locations throughout California, and found that populations behave in different ways. Some populations exhibit large fluctuations in size while others remain relatively stable. Our research suggests that the degree to which individuals within a population vary has a big effect on the dynamics of populations. We found that populations with low genetic diversity exhibited stronger competition and less fluctuation in size than populations with high genetic diversity. A main reason for this is that low-diversity populations exhibited more spatial clustering, which likely lead to more intense competition. We are continuing to study this phenomenon and it seems like the genetic and phenotypic composition of populations may play a large role in governing their dynamics.
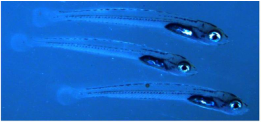 13-day old larvae of California grunion. Lab studies confirm that low pH increases mortality and reduces growth rates
13-day old larvae of California grunion. Lab studies confirm that low pH increases mortality and reduces growth rates
Fish in hot & sour soup: Can our piscine friends survive changes in ocean temperature and chemistry?
As carbon dioxide has accumulated in the atmosphere, the temperature of the globe and its oceans has risen. Excess CO2 in the atmosphere diffuses into the ocean where it disrupts ocean chemistry and reduces the average pH of seawater -- a phenomenon known as ocean acidification. If carbon emissions continue to increase, the ocean will become both hotter, and more sour (as ocean pH becomes more acidic).
It can be stressful and difficult for fish larvae to regulate their internal pH when the pH of the seawater around them changes. These stresses can cause changes in behavior, growth, and mortality rates. But is it all bad news?
Maybe not. If certain genotypes in the population are naturally more tolerant of low pH seawater, then populations may evolve to become more resistant to the effects of ocean acidification. The key is understanding how much genetic variation exists within populations. If there is a lot, then populations may evolve quickly. If there is little, then populations may evolve slowly (if at all), and populations will suffer the ill effects of ocean acidification for a long time.
As carbon dioxide has accumulated in the atmosphere, the temperature of the globe and its oceans has risen. Excess CO2 in the atmosphere diffuses into the ocean where it disrupts ocean chemistry and reduces the average pH of seawater -- a phenomenon known as ocean acidification. If carbon emissions continue to increase, the ocean will become both hotter, and more sour (as ocean pH becomes more acidic).
It can be stressful and difficult for fish larvae to regulate their internal pH when the pH of the seawater around them changes. These stresses can cause changes in behavior, growth, and mortality rates. But is it all bad news?
Maybe not. If certain genotypes in the population are naturally more tolerant of low pH seawater, then populations may evolve to become more resistant to the effects of ocean acidification. The key is understanding how much genetic variation exists within populations. If there is a lot, then populations may evolve quickly. If there is little, then populations may evolve slowly (if at all), and populations will suffer the ill effects of ocean acidification for a long time.
 Grunion run!
Grunion run!
Using California Grunion as a model species, we are measuring the amount of genetic variation underlying larval tolerance to low-pH seawater. Grunion are an important species in Southern California, and are best known for their habit of spawning on sandy beaches! This makes them a great model species because when the time is right, you can run down to the seashore, grab a few grunion and make them spawn into plastic cups. Doing this allows you to create groups of larvae that are close relatives (e.g., half and full sibs) and groups that are unrelated. The degree to which larval tolerance of low-pH seawater is similar for closely related larvae indicates the amount of genetic variation underlying pH tolerance.
Rovers Return: Tracking larval dispersal and population connectivity within a marine fish metapopulation
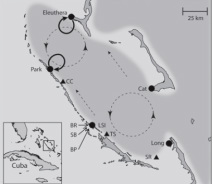
Our study area, Exuma Sound, Bahamas
Many marine species have an adults that live near the seafloor and larvae that develop within the plankton. Larvae often spend weeks to months in the water column and even if adults to not move much, larvae have the potential to disperse long distances. Where larvae go matters, and it is important to know how many larvae come home and how many go to other populations. In addition, the health of the regional population (the metapopulation) will depend on the degree to which local populations of adults are connected by the movement of larvae. Together with colleagues Mark Hixon (Univ. of Hawaii), Mark Christie (Purdue Univ.), Chris Stallings (Univ. of South Florida) and Tim Pusack (Williams College), we have been studying connections between demography and dispersal among populations of damselfish at four widely spaced islands in the Bahamas. By combining direct measures of offspring production with genetic parentage analysis, we could estimate how many genetically "tagged" larvae were produced at a particular island, and how many of those fish returned to study island as juveniles. Our results suggested that 13% of recruits returned to their natal islands, on average, and that exchange of larvae was appreciable for islands up to 129 km apart. Details can be found in our recent publication (Johnson et al. 2018, Ecology) and in our previous publications (Christie et al. 2010, Molecular Ecology; Pusack et al. 2014, Molecular Ecology).
Quantifying Evolutionary Potential of Marine Fish Larvae
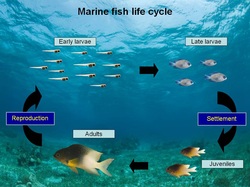
For your average marine fish larva, life is hard. There are many predators that want to eat you, and finding food and habitat can be difficult. The result is that many larvae are produced, but very few survive (it is common for < 1% of to make it through the larval stage).
And life is not fair. Larvae with certain qualities (e.g., larger size, better swimming ability) have a better chance of surviving. This combination of intense, selective mortality suggests that marine fish larvae have the potential to evolve very quickly. However, like many biological phenomena, things are not so simple.

Checking a damselfish nest for eggs
Responses to selection also depend on the extent of genetic variability underlying traits that are selected, as well as selection that occurs at other times in the life cycle. Together with colleagues at Oregon State University, I have extensively studied a model species (bicolor damselfish) and estimated the genetic, environmental, and maternal sources of variation in larval size and swimming performance --two traits that strongly affect survival. Although there was substantial genetic variation in larval traits, we found that the evolutionary potential of larvae was actually quite low because selection favoring larger larvae was largely balanced by selection favoring mothers who tend to produce many, small offspring (Johnson et al. 2010 Evolution).
A small fry's perspective on fisheries-induced evolution
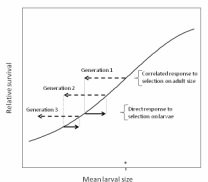
Many fisheries selectively remove large individuals from the population. This may lead to rapid evolution of fished populations, though the potential for evolutionary responses is not always clear. For example, even if selection acts on a single trait (adult body size), other traits that are genetically correlated may also evolve.
My studies of damselfish larvae found substantial genetic correlations between adult size and the size and swimming performance of larvae. The implication is that selection on adult size (as occurs in many fisheries) can cause a substantial genetic response of traits that affect larval survival. Moreover, because the link between larval traits and survival is strong, a correlated, evolutionary reduction in larval size can result in a significant decline in population replenishment (Johnson et al. 2011 Evolutionary Applications). Some of my current work uses population projection models to examine how correlated responses to selection during various stages of the life cycle may influence fisheries-induced evolution, and how these reciprocal evolutionary effects ultimately regulate population dynamics.
My studies of damselfish larvae found substantial genetic correlations between adult size and the size and swimming performance of larvae. The implication is that selection on adult size (as occurs in many fisheries) can cause a substantial genetic response of traits that affect larval survival. Moreover, because the link between larval traits and survival is strong, a correlated, evolutionary reduction in larval size can result in a significant decline in population replenishment (Johnson et al. 2011 Evolutionary Applications). Some of my current work uses population projection models to examine how correlated responses to selection during various stages of the life cycle may influence fisheries-induced evolution, and how these reciprocal evolutionary effects ultimately regulate population dynamics.
Long, strong, and down to get the friction on?

Sperm from Galeolaria caespitosa
Many marine species are broadcast spawners. That is, they reproduce by releasing eggs and sperm into the water column. Also, many broadcast spawners can't move, or don't move much as adults. For these species, an individual's fertilization success (and subsequent fitness) depends heavily on the characteristics of their eggs or sperm.
Together with Dustin Marshall of Monash University, we have been examining how certain morphological characteristics of sperm can influence fertilization success in Galeolaria caespitosa, a tube worm found in the rocky intertidal of southeast Australia. Morphology can influence the fertilization success of sperm by affecting a sperm's ability to swim (and contact eggs before other sperm do). However, the swimming speed of sperm has to be considered in the right environmental context. Because of their small size, sperm move through a fluid environment that is dominated by viscosity, rather than inertia (picture yourself doing a front crawl through a pool of molasses to get a rough idea of what it's like). In this environment, swimming speed of sperm is determined by a balance between thrust, which is proportional to tail length, and drag, which is proportional to head size. Although sperm with smaller heads should have the capability to swim faster, larger heads may be associated with greater energy reserves. By measuring sperm morphology and fertilization success, we're testing whether it is better to be long (and strong), and whether it's beneficial to produce sperm with larger heads, even though this may lead to greater frictional drag which can constrain swimming speed (Johnson et al. 2013, Evolution).
Together with Dustin Marshall of Monash University, we have been examining how certain morphological characteristics of sperm can influence fertilization success in Galeolaria caespitosa, a tube worm found in the rocky intertidal of southeast Australia. Morphology can influence the fertilization success of sperm by affecting a sperm's ability to swim (and contact eggs before other sperm do). However, the swimming speed of sperm has to be considered in the right environmental context. Because of their small size, sperm move through a fluid environment that is dominated by viscosity, rather than inertia (picture yourself doing a front crawl through a pool of molasses to get a rough idea of what it's like). In this environment, swimming speed of sperm is determined by a balance between thrust, which is proportional to tail length, and drag, which is proportional to head size. Although sperm with smaller heads should have the capability to swim faster, larger heads may be associated with greater energy reserves. By measuring sperm morphology and fertilization success, we're testing whether it is better to be long (and strong), and whether it's beneficial to produce sperm with larger heads, even though this may lead to greater frictional drag which can constrain swimming speed (Johnson et al. 2013, Evolution).
